![]()
Perfil metabólico de cordeiros com diferentes níveis do ALL-G RichTM na dieta
3 - Universidade Federal de Viçosa
4 - Universidade Federal de Uberlândia
5 - Universidade Federal de Uberlândia
6 - Universidade Federal de Uberlândia
7 - Universidade Federal de Uberlândia
8 - Universidade Federal do Tocantins
RESUMO -
Objetivou-se avaliar o efeito de diferentes níveis de ácido docosahexaenóico (DHA, produto comercial ALL-G RichTM) no concentrado sobre o metabolismo proteico, energético, hepático e mineral. O experimento foi realizado no setor de caprinos e ovinos na fazenda experimental Capim Branco em Uberlândia – MG. Foram utilizados 24 cordeiros (12 machos e 12 fêmeas) mestiços (Dorper x Santa Inês), mantidos confinados por quarenta dias recebendo dieta com relação volumoso concentrado de 18:82% com níveis de 0%, 3%, 6% e 9% no concentrado de ácido docosahexaenóico (DHA, produto comercial ALL-G RichTM). O delineamento experimental foi inteiramente ao acaso, em arranjo fatorial com medidas repetidas no tempo. Não houve diferença estatística para o perfil glicêmicos e metabólicos avaliados em relação aos tratamentos e classe sexual. O comportamento das equações de regressão em relação aos períodos é relacionado á fase de crescimento do animal, identificando maior atividade hepática. Conclui-se que o DHA pode ser utilizado para cordeiros tanto machos como fêmeas em recria.
Metabolic Profile of Lambs with Different ALL-G RichTM Levels in Diet
ABSTRACT - The objective of this study was to evaluate the effect of different levels of docosahexaenoic acid (DHA, commercial product ALL-G RichTM) in the concentrate on protein, energy, hepatic and mineral metabolism. The experiment was carried out in the goats and sheep sector at the Capim Branco experimental farm in Uberlândia - MG. Twenty-four lambs (12 males and 12 females) crossbreed (Dorper x Santa Inês) were kept confined for forty days receiving a diet with a concentrated ratio of 18: 82% with levels of 0%, 3%, 6% and 9% Docosahexaenoic acid concentrate (DHA, commercial product ALL-G RichTM). The experimental design was completely randomized with measures repeated over time. There was no statistical difference for the glycemic and metabolic profile evaluated in relation to treatments and sexual class. The behavior of the regression equations in relation to the periods is related to the growth phase of the animal, identifying higher hepatic activity. It is concluded that DHA can be used for both male and female lambs in rearing.Introdução
A alimentação geralmente representa o maior custo dentro dos sistemas de produção animal (Almeida, 2005). Visto essa importância, é fundamental um planejamento nutricional no qual seja possível equalizar os custos de produção da alimentação dos animais e o desempenho dos mesmos que irá representar a receita final do sistema de produção. Assim é fundamental ao nutricionista responsável o conhecimento de diferentes estratégias nutricionais. Os diferentes tipos de gordura entram nesse contexto como uma opção devido ao alto teor energético. Novas fontes como gorduras provenientes de algas podem ser uma boa opção, visto seu diferente perfil nutricional principalmente por constituir fontes de ácidos graxos como o do ácido docosahexaenóico (DHA, ômega 3). Muitos são os benefícios sobre a utilização de ácidos graxos da série ômega 3 na alimentação humana, como o melhor desempenho do cérebro (Almeida e Bueno Franco, 2006), além do desenvolvimento da visão, cognição e imunidade. A utilização de fontes de ômega 3 na alimentação animal poderá gerar produtos, como carne e leite ricos nesse tipo de gordura e com isso trazer benefícios à saúde humana. Já é possível encontrar no mercado alguns produtos de origem animal como ovos enriquecidos com PUFA (Poly Insaturated Fatty Acid do tipo Ômega 3). Maiores quantidades de lipídeos não protegidos e poli-insaturados (como o DHA) na dieta podem afetar a colonização, a capacidade fermentativa da fibra pelos micro-organismos do rúmen e a gordura do leite. Sendo assim é necessário estudar como os níveis de suplementação desses produtos na alimentação animal podem afetar a dinâmica ruminal e o metabolismo. Objetivou-se então avaliar o efeito de diferentes níveis de ácido docosahexaenóico (DHA, produto comercial ALL-G RichTM) no concentrado sobre o metabolismo proteico, energético, hepático e mineral de ovinos.Revisão Bibliográfica
Os animais ruminantes possuem características anatômicas e simbióticas capazes de utilizar eficientemente carboidratos estruturais e carboidratos solúveis como fonte de energia. A fermentação nestes animais é obtida por atividades físicas, químicas e microbiológicas, convertendo os componentes dietéticos a ácidos graxos voláteis, proteína microbiana e algumas vitaminas (Berchielli, Pires e Oliveira, 2006), constituindo os elementos básicos produzidos pela fermentação que são aproveitados pelo animal. Diversas fontes lipídicas são usadas na nutrição de ruminantes. Cada tipo de alimento rico em gordura como soja grão, o caroço de algodão, a canola, a linhaça, óleo de soja, óleo de girassol, óleo de algodão, a gordura inerte de óleo de palma, gordura inerte de óleo de soja e recentemente as algas marinhas, possuem suas próprias particularidades e limites de uso, portanto devem ser estudados de modo a não prejudicar o funcionamento ruminal. Dentre essas se têm os ácidos graxos ômega-3 (ou n-3) são caracterizados por constituir insaturações. A designação de ômega tem relação com a posição da primeira dupla ligação, contando a partir do grupo metílico final da molécula do ácido graxo, sendo a primeira dupla ligação entre o terceiro e o quarto átomo de carbono. Os ácidos alfa-linolênico (18:3 n-3), eicosapentaenoico (20:5 n-3) e docosahexaenóico (22:6 n-3) são exemplos de ácidos graxos da série ômega 3. As bactérias do rúmen possuem certa ineficiência em digerir a fibra da ração na presença de lipídeos que geralmente se encontram na forma de triglicerídeos (ácidos graxos + glicerol). Tal fato irá depender da quantidade, da natureza do lipídeo (poli-insaturados ou monossaturados são mais tóxicos) e da fonte de fibra, porém a fermentação do amido não é afetada (Valadares Filho e Pina, 2006). Um mecanismo realizado como resposta é a biohidrogenação dos ácidos graxos na forma não esterificada, convertendo esses em ácidos graxos saturados, por sua vez menos tóxicos. Para isso ocorre a quebra do triglicerídeo dentro do rúmen, onde o glicerol é rapidamente fermentado e convertido a ácido graxo volátil e o ácido graxo de cadeia longa ou média sofre uma isomerização da forma cis para a forma trans e imediatamente ocorre a hidrogenação do composto se esse for insaturado (Palmquist e Mattos, 2006). Há algum tempo foram desenvolvidos e lançados no mercado, gorduras inertes para alimentação de ruminantes. Essas gorduras geralmente são envolvidas por sabões de cálcio que conferem proteção da degradação ruminal. Esses sabões são estáveis em pH próximo a neutralidade (pH ruminal), porém se desfazem com o pH ácido do abomaso liberando os lipídeos para a digestão no intestino delgado. Essa proteção retira o efeito deletério do lipídeo sobre os micro-organismos do rúmen, constituindo uma boa estratégia na alimentação de ruminantes, visto o alto teor energético da molécula lipídica. É importante ressaltar que a ocorrência de acidose ruminal culminará na dissolução dos sabões de cálcio dentro do rúmen, liberará os lipídeos promovendo o efeito deletério sobre as bactérias e a fibra com capacidade de diminuir a fermentação ruminal e a gordura do leite. O uso de gordura inerte requer o controle de pH do rúmen que pode ser feito via dieta onde a adição de agentes tamponantes na ração são uma boa opção. Os óleos de palma e soja são exemplos de gorduras inertes. Geralmente o que mais limita o uso desses produtos é o seu alto valor de mercado. Já fontes de gorduras não protegidas apresentam menor custo, porém seu consumo acaba por ser limitado há quantidades menores que 10% da matéria seca, devido ao prejuízo sobre o ambiente ruminal. Existem na literatura diversos trabalhos trazendo diferentes resultados com o uso das mais diversos tipos de alimentos ricos em lipídios e suas combinações na nutrição. Os alimentos que contem lipídeos provindos de algas marinhas ricas em ácidos graxos poli-insaturados como o DHA merecem atenção já que são produtos novos que podem trazer inúmeros benefícios à saúde humana. Porém, ainda não se conhece seus níveis seguros na alimentação dos animais e sua influência sobre o metabolismo.Materiais e Métodos
O presente trabalho foi conduzido entre 11 de janeiro a 19 de fevereiro de 2015, na Fazenda experimental Capim Branco, Setor de Ovino e Caprinocultura da Universidade Federal de Uberlândia, localizada na região sudoeste de Minas Gerais – Brasil. Foram utilizados 24 ovinos mestiços (Dorper x Santa Inês) 12 machos e 12 fêmeas recém-desmamados com idade média 90 dias e peso corporal médio de 17 kg. Esses foram alojados em quatro baias coletivas sendo seis animais por baia (três machos e três fêmeas), com piso ripado, comedouro, saleiro e bebedouro. Os animais foram casqueados e vermifugados com cloridrato de levamisol no período de adaptação, dez dias antes do início do experimento. Além disso, foi realizado controle da eimeriose com Sulfaquinoxalina®. Ao longo do experimento foram realizados exames para verificação da verminose e eimeriose. A ração utilizada foi composta por silagem de milho, farelo de milho, farelo de soja, mineral específico para espécie e ALL-G RichTM (Tabela 1), sendo balanceada para atender as necessidades nutricionais de ganho 200g/animal/dia segundo o NRC (2007). O arraçoamento foi dividido em dois períodos, sendo fornecido às 8:00 e às 16:00 horas. A relação volumoso:concentrado adotada foi de 18:82% (na matéria seca), respectivamente. Os tratamentos foram inclusão de 0%, 3%, 6 % e 9 % de ácido docosahexaenóico (DHA, provindo do produto comercial ALL-G RichTM) no concentrado ofertados diariamente. Para os períodos experimentais obteve-se intervalo de 15 dias entre avaliações, sendo divididos em 0, 15, 30 e 40 dias. A cada novo período experimental os animais foram pesados para ajuste do fornecimento de ração, conforme recomendação do NRC (2007). A glicemia foi realizada no início e fim do experimento, sendo as coletas ás 8, 11, 14, 17 e 20 horas em um dia. Para colheita do sangue puncionou-se a veia jugular com auxílio de um vacuntainer acoplado a tubo contendo fluoreto mais EDTA. Após esse procedimento, as amostras foram centrifugadas e pipetadas em mini tubos para congelamento. As mesmas foram analisadas no laboratório de patologia clínica da Universidade Federal de Uberlândia, com aparelho automatizado e utilizando o kit comercial da LabTest®. A coleta das 8 horas foi realizada com os animais em jejum, após forneceu-se o primeiro trato e então realizou as demais coletas do dia e após a coleta das 20 horas ofertou-se o segundo trato. As coletas de sangue foram realizadas durante os períodos experimentais no período da manhã antes da primeira alimentação, por venopunção jugular com auxílio de vacuntainner e tubos sem anticoagulante. Logo após, as amostras de cada animal foram centrifugadas e pipetadas em eppendorfs para posterior análise laboratorial usando kit comercial da Lab Test®. Os metabólitos energéticos determinados foram: colesterol (mg/dL), triglicerídeos (mg/dL), VLDL (mg/dL, obtida pela divisão dos valores de triglicerídeos por cinco) e frutosamina (μmol/L). Os metabólicos hepáticos foram: aspartato aminotrasferase (U/L), gama glutamil tranferase (U/L) e fosfatase alcalina (U/L). Os metabólicos proteicos foram: albumina (g/dL), proteína total (g/dL), creatina (mg/dL), ureia (mg/dL) e ácido úrico (mg/dL). E por fim, os metabólicos minerais foram: Fósforo (mg/dL), Cálcio (mg/dL), relação Cálcio/Fósforo e Magnésio (mg/dL). O delineamento utilizado foi inteiramente casualizado (DIC) em arranjo fatorial (duas classes sexuais e quatro níveis de inclusa) com medidas repetidas ao tempo. Para análise de glicemia analisou-se efeito de medida repetida no tempo para os períodos (equação de regressão) e medida repetida no tempo para os horários de coleta. Para comparação dos tratamentos e períodos utilizou-se o estudo de regressão (P<0,05). Já para as médias relativas ao sexo foi feito o teste SNK (P<0,05). Todas as analises foram realizadas em software SAS 9.4.Resultados e Discussão
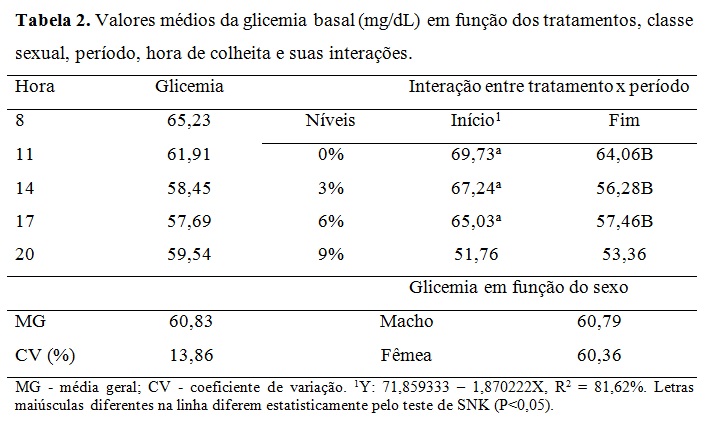
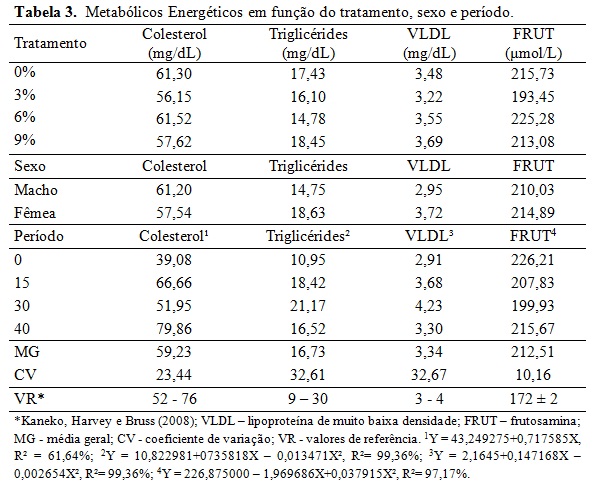
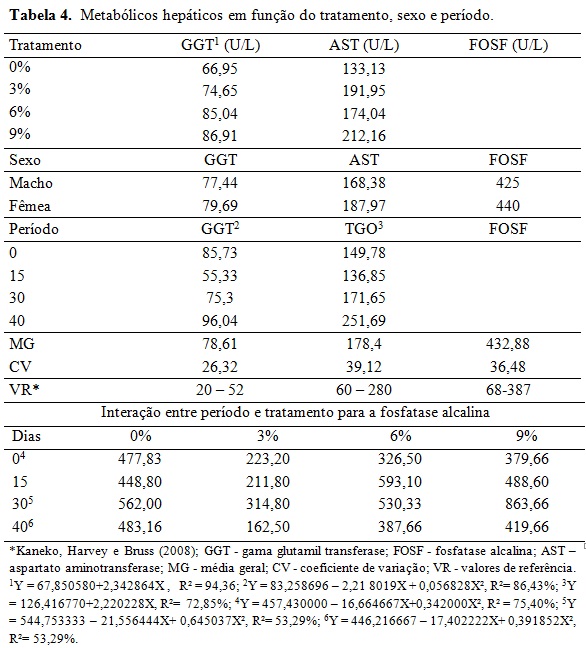
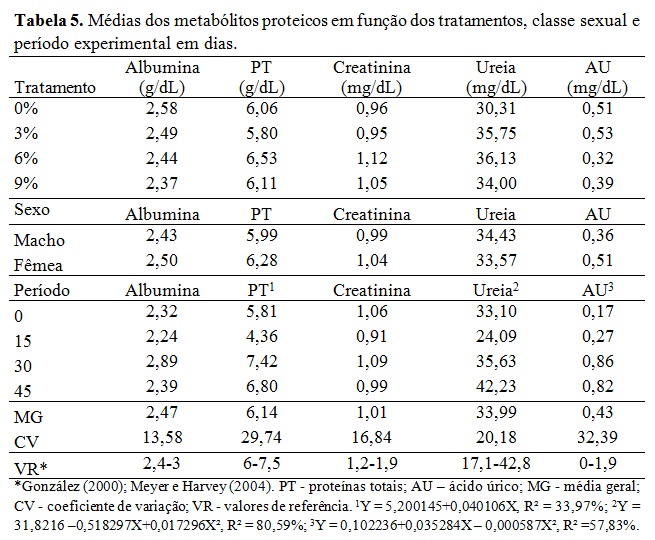
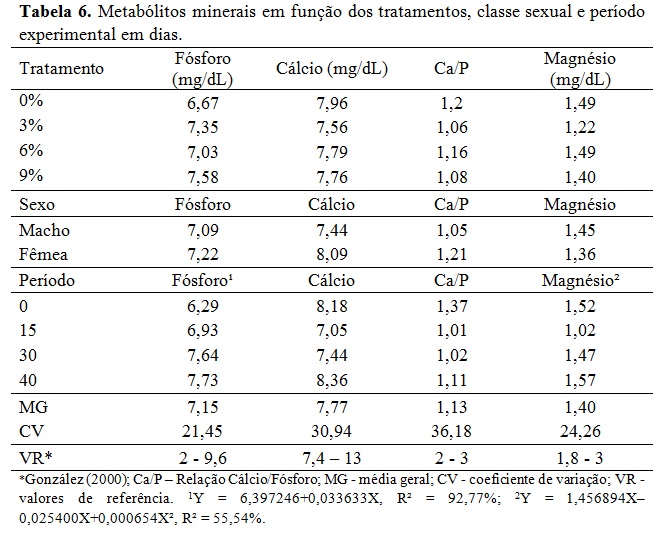
Não houve efeito da classe sexual e dos momentos de colheita ao longo dia (Tabela 2). A interação dos níveis glicêmicos foram maiores no início do experimento quando comparados ao final para os tratamentos 0%, 3% e 6%. Isso pode ser explicado pela idade dos animais e o pôs desmame, portanto antes do período experimental tinham uma alimentação rica em açúcares e ao final com idade próxima a puberdade, ocorreu à redução dos níveis basais de glicose, uma vez que esta passa a ser formada indiretamente (gliconeogênese). Nos tratamentos 3% e 6% o lipídeo reduziu consideravelmente os níveis glicêmicos ao final em comparação ao tratamento 0%, assim como no tratamento 9% o teor lipídico diminuiu níveis glicêmicos ao início e ao final, porém não sem diferença estatística. Assim como observado por Silva et al., (2010) nos níveis sanguíneos de glicose em cabras leiteiras alimentadas com diferentes fontes lipídicas e os valores de glicemia desses autores ficaram semelhantes ao valor do tratamento A desse estudo. Os valores adequam-se ao valor de glicemia basal (50 a 80 mg/dL) em ovinos citado por Kaneko, Harvey e Bruss (2008). A equação de regressão da Tabela 2 mostrou que à medida que aumentou os níveis de gordura na ração houve decréscimo linear dos níveis basais de glicose na fase inicial do experimento, reflexo da adaptação dos animais a dieta após o início do período experimental. Embora os níveis glicêmicos tenham caído nos tratamentos de maior teor de gordura, não se pode entender que o metabolismo hepático e energético desses animais era menos intenso, o que se pode esperar é a mudança de rota metabólica da energia. Pode-se observar que a frutosamina (Tabela 3) encontrava-se em todos os tratamentos bem acima do valor de referência descrito por Kaneko, Harvey e Bruss (2008) o que evidência alta energia nos animais. Ressalta-se que essas tabelas internacionais possam não refletir as condições metabólicas dos animais criados no sistema brasileiro. A frutosamina é um metabólico que representa a associação da glicose com a albumina. A taxa de renovação desse metabólito fica em torno de 70 dias, a concentração elevada reflete também o alto metabolismo energético, o que pode sugerir alta atividade hepática, sem querer refletir em desordem nesse órgão. Os níveis de colesterol e VLDL não tiveram efeito sobre os diferentes tratamentos e classe sexual e mantiveram-se dentro dos valores de referência. Segundo González et al., (2002) o colesterol sanguíneo pode ser de origem endógena sintetizado a partir da acetil-CoA ou exógena, provenientes dos alimentos, principalmente do amido. Além disso, o colesterol é precursor de hormônios esteróides ligados à reprodução. O fato dos animais estarem em fase de crescimento, próximos a puberdade, contribuiu para aumentar essas variáveis no período, além da própria característica da ração que continham alta energia em amido e lipídeo promovendo o aumento do estoque de energia corporal nos animais. Os triglicerídeos, VLDL (lipoproteína de muito baixa densidade), LDL (lipoproteína de baixa densidade) e HDL (lipoproteína de alta densidade) mantiveram-se dentro dos valores de referência, com incremento durante o período, mas com queda no final. Silva et al., (2010) encontraram aumento dos triglicerídeos sanguíneos com a inclusão de lipídeos na ração em comparação a dieta que não recebeu adição lipídica de caroço de algodão ou faveleira em cabras leiteiras. Porém as fontes lipídicas desses autores possuem uma natureza lipídica diferente do ácido docosaexahenóico. Em relação às enzimas hepáticas, houve efeito da inclusão do DHA somente para a gama glutamil tranferase (GGT) sendo o padrão da resposta linear em relação ao aumento do nível de DHA na dieta (Tabela 4). No período houve, de forma geral, acréscimo dos níveis dos metabólicos hepáticos. Pode-se entender que com o crescimento corporal dos animais, houve aumento do seu metabolismo devido à deposição muscular, início da deposição lipídica na carcaça e aumento da carga hepática devido ao metabolismo do lipídeo. Não houve efeito de sexo em nenhum metabólico analisado, indicando comportamento semelhante no perfil metabólico de machos e fêmeas. A gama glutamil transferase (GGT) é uma enzima usada como parâmetro bioquímico para indicar alterações hepáticas e biliares. Em todos os tratamentos essa enzima está acima dos valores de referência descritos por Kaneko, Harvey e Bruss (2008), indicando que o fígado desses animais encontrava-se em alto metabolismo. Os valores de aspartato aminotransferase (AST) estão dentro da faixa de normalidade segundo Kaneko, Harvey e Bruss (2008), porém próximos ao limite superior principalmente ao final do experimento e nos tratamentos de maior teor lipídico reiterando sobrecarga hepática desses animais com o decorrer de uso da fonte lipídica. Houve efeito de interação entre tratamento e período para a fosfatase alcalina (FOSF) nos dias 0, 30 e 45 (Tabela 4) atentando-se ao alto valor da enzima nos animais do tratamento 9% no período de 30 dias, esta enzima é também utilizada para condições patológicas, como a sobrecarga hepática. Para o perfil proteico não houve efeito significativo sobre a inclusão de DHA nos diferentes tratamentos ou em função da classe sexual (Tabela 5). A ureia sanguínea reflete diretamente a amônia gerada no rumen pela degradação dos compostos nitrogenados e que não foi convertida em proteína microbiana. Essa amônia excedente atravessa a parede ruminal, indo diretamente para ser transformada em ureia no fígado com alto gasto energético. Essa ureia então pode ser eliminada na urina, retornar ao rumen via saliva ou difusão na parede ruminal e ser eliminada no leite em caso de animal em lactação (Wittwer, 2000). A Tabela 4 mostra variação da ureia no período indicando maior escape de amônia do rumen, principalmente ao final sugerindo falta de sinergismo entre a relação energia proteína. Além disso, pode refletir na incapacidade da microbiota fermentar as fontes de carboidratos em função dos efeitos deletérios desse lipídeo. Ocasionando assim aumento da ureia ao longo do experimento. A proteína total apresentou uma resposta linear positiva no período experimental. A albumina e a proteína total refletem no status nutricional do animal e na quantidade de proteína da dieta (González, 2000), então baixos níveis circulantes desses metabólicos podem refletir deficiência proteica na ração. Os valores desses metabólicos encontram-se dentro da faixa normal ou bem próximo dela refletindo correta quantidade de proteína fornecida aos animais que mantiveram bom status nutricional. Os valores de creatinina ficaram dentro da faixa normal e não houve diferença estatística entre tratamentos e no período mostrando que a função renal dos animais do experimento não estava comprometida. Os efeitos acima para proteína total, albumina e creatinina estão dentro de uma resposta esperada para a espécie (González, 2000; Meyer e Harvey, 2004). Vale ressaltar que no período de 15 dias todos metabólicos proteicos, com exceção do ácido úrico, estavam com valores mais reduzidos mostrando uma fase de adaptação dos animais à dieta e a conviver sem a companhia de suas mães. Os metabólicos minerais analisados não apresentaram diferença significativa entre os tratamentos (Tabela 6). Também não houve diferença dos metabólitos minerais em relação à classe sexual, indicando comportamento metabólico semelhante de machos e fêmeas que entraram no experimento por volta de 90 dias e saíram próximos a puberdade. O fósforo possui importantes funções fisiológicas como componente dos ossos, dos fosfolipídios e da molécula de ATP. Os seus níveis são aumentados quando a dieta possui alto concentrado provindo dos grãos. O fósforo é intimamente ligado ao cálcio, porém possui um menor controle endócrino que este, sendo comuns maiores variações nas suas concentrações sanguíneas. Avaliando o período, foi possível observar aumento dos níveis de todos os metabólitos minerais demonstrando crescimento dos animais, entretanto, foi verificada diferença significativa apenas nos valores de fósforo e magnésio. O aumento linear do fósforo que pode ser explicado pelo fato dos animais estarem em fase de crescimento onde o cálcio é requerido para crescimento dos ossos como é possível observar pela baixa relação cálcio/fósforo (desejável 2:1) (González, 2000), o que indica rápido crescimento e ganho de massa corporal. O cálcio é intimamente ligado ao metabolismo possuindo diversas funções como componente estrutural além de participar da contração muscular. Seus níveis geralmente não são muito variáveis permanecendo em uma estreita faixa (González, 2000). Por esse fato o cálcio é menos eficiente para refletir o estado nutricional comparado ao fósforo e ao magnésio. Não se observou diferença significativa e seus níveis sanguíneos que estavam próximos ao limite inferior dos valores de referencia indicando o crescimento dos animais com a deposição óssea. Os teores de magnésio apresentaram padrão quadrático e abaixo dos valores de referência. Isso pode ser explicado pelo fato dos animais estarem ingerindo uma fonte de gordura em diferentes níveis, na qual o magnésio possui função metabólica importante. Outro fator pode ser o fato dos animais estarem em fase de crescimento corporal, o que inclui deposição desse mineral na estrutura óssea. É importante ressaltar que, por não existir um controle homeostático rigoroso do magnésio, seus níveis estão relacionados ao seu fornecimento através da alimentação. Logo, uma dieta com baixos teores de magnésio deve ser ajustada para atender às necessidades dos animais.Conclusões
O aumento da inclusão de DHA provindo do produto comercial ALL G Rich® no concentrado elevou os níveis das enzimas hepáticas dos animais, porém não afetou as demais vias metabólicas, podendo ser utilizado na alimentação de borregos em confinamento na fase de recria.Gráficos e Tabelas