![]()
BALANÇO ELETROLÍTICO TOTAL (BET) PARA CODORNAS JAPONESAS (COTURNIX COTURNIX JAPONICA)
2 - Instituto Federal de Educação, Ciência e Tecnologia do Norte de Minas Gerais-IFNMG
3 - Universidade Federal de Minas Gerais-UFMG
4 - Instituto Federal de Educação, Ciência e Tecnologia do Norte de Minas Gerais-IFNMG
5 - Instituto Federal de Educação, Ciência e Tecnologia do Norte de Minas Gerais-IFNMG
6 - Instituto Federal de Educação, Ciência e Tecnologia do Norte de Minas Gerais-IFNMG
7 - Instituto Federal de Educação, Ciência e Tecnologia do Norte de Minas Gerais-IFNMG
8 - Instituto Federal de Educação, Ciência e Tecnologia do Norte de Minas Gerais-IFNMG
RESUMO -
Com o intuito de avaliar o melhor valor de balanço eletrolítico total (BET) 240 codornas japonesas, durante o período de 25 a 85 dias de vida, foram distribuídas em um delineamento inteiramente casualizados, num total de cinco tratamentos (750, 1000, 1250, 1500 e 1750 mEq/kg), com oito repetições e seis aves por unidade experimental. As características de desempenho analisadas foram as seguintes variáveis: postura (%), peso médio dos ovos (g), consumo de ração (g/ave/dia), conversão alimentar por massa de ovos (g de ração/g de ovo), conversão alimentar por dúzia de ovos (g de ração/dz de ovo) e uniformidade (%). As características qualitativas dos ovos foram: os pesos de gema, albúmen e casca, além disso, foram feitas avaliações de cor e densidade dos ovos. Os diferentes BET foram capazes de influenciar o desempenho das aves. O valor de balanço eletrolítico que propiciou menor consumo sob mesmo desempenho foi BET=1359 μeq/kg, e o que resultou em melhor conversão alimentar por dúzia de ovos e uniformidade foi de BET = 1000 μeq/kg. Os parâmetros de densidade, coloração da gema, unidade Haugh, peso da gema, peso da casca e peso do ovo não foram afetados pelos BET experimentais. Os valores de peso do albúmen foram afetados pelos diferentes BET. O valor de BET que propiciou peso do albúmen máximo, descrito segundo a equação, é de BET= 1600 em μeq/kg de cargas nas rações.
TOTAL ELECTROLYTE BALANCE (TEB) FOR JAPANESE QUOTES (COTURNIX COTURNIX JAPONICA)
ABSTRACT - Aiming to evaluate the best total electrolytic balance (TEB) value, a total of 240 Japanese quails were used during a period of 25 to 85 days, with a total of five treatments (750, 1000, 1250, 1500 and 1750 mEq / kg) distributed in a completely randomized design with eight replicates and six birds per experimental unit. The performance characteristics analyzed were the following variables: laying (%), mean egg weight (g), feed intake (g / bird / day), feed conversion per egg mass (g feed / g egg), feed conversion per dozen eggs (g ration / egg dz) and uniformity (%). The egg’s qualitative characteristics evaluated were: weights of yolk, albumen and shell; in addition, egg color and density were evaluated. The different BETs were able to influence the performance of the birds. The TEB value that provided the lowest consumption under the same performance was TEB = 1359 μeq / kg, which resulted in better feed conversion per dozen eggs and uniformity was BET = 1000 μeq / kg. The parameters of density, yolk color, Haugh unit, yolk weight, shell weight and egg weight were not affected by the experimental TEB. The albumen weights were affected by different TEB. The TEB value of the maximum albumen, described by the equation, is the TEB = 1600 in μeq /kg of feed in the feed.Introdução
Os eletrólitos da ração consumida pelos animais exercem influência no equilíbrio ácido-base e, consequentemente, afetam processos metabólicos relacionados ao crescimento, à resistência a doenças, à sobrevivência ao estresse e aos parâmetros de desempenho (LIN et al., 2006). A manutenção do equilíbrio ácido-base do organismo animal tem grande importância fisiológica e bioquímica, uma vez que as atividades das enzimas celulares, trocas eletrolíticas e manutenção do estado estrutural das proteínas dos organismos são profundamente influenciadas por pequenas alterações no pH sanguíneo (BORGES et al., 2007). Apesar dos limites inferior e superior de pH necessários à manutenção da vida serem, respectivamente, 6,8 e 7,8 (VIEIRA et al., 1979), os valores normais de pH sanguíneo devem encontrar-se entre os limites de 7,35 e 7,45, para o correto funcionamento das moléculas bioativas do organismo. O distúrbio ácido-base primário é normalmente indicado pela determinação do pH do sangue e dos componentes respiratório (PCO2) e metabólico (HCO3-), conforme descrito por Ahmad & Sarwar (2006). O dióxido de carbono total (CO2 total), excesso de bases (EB) e a pressão parcial de O2 (PO2) também têm sido apontados como indicadores da condição ácido-base do sangue (VIEIRA et al., 1979). Considerando que as enzimas celulares, as trocas eletrolíticas e a manutenção do estado estrutural das proteínas são diretamente afetadas por pequenas variações no pH sanguíneo, o controle da variação do pH fisiológico se torna essencial para o perfeito funcionamento dos processos bioquímicos (MACARI et al., 2002). Mongin (1981) estudou os fundamentos do balanço eletrolítico (BE) para aves e suínos e concluiu que pode-se descrever o equilíbrio entre os íons por um cálculo envolvendo os principais deles, cuja fórmula representativa é o resultado da soma dos íons Na+ e K+, subtraindo-se o íon negativo Cl-, denominado BE. Porém, as dietas atuais são repletas de ingredientes capazes de influenciar no balanço eletrolítico, porém estes não são contabilizados no modelo proposto por Mongin (1981), como os aminoácidos sintéticos por exemplo. Araújo et al. (2011) propôs um novo cálculo de balanço eletrolítico onde estes eletrólitos fossem contabilizados. Assim, com o avanço do conhecimento sobre estes eletrólitos, e a influência de seus mEq sobre o metabolismo das aves, torna-se necessário contabilizá-los dentro do cálculo de balanço eletrolítico. Assim este trabalho foi realizado com o intuito de avaliar o melhor valor de balanço eletrolítico total (BET) para codornas japonesas, durante o período de 25 a 85 dias de vida.Revisão Bibliográfica
A composição mineral da ração afeta o equilíbrio ácido-base e vários aspectos do desempenho animal (MURAKAMI, 2000). Assim manutenção do equilíbrio ácido-base do meio interno tem grande importância fisiológica e bioquímica, uma vez que as atividades das enzimas celulares, trocas eletrolíticas e manutenção do estado estrutural das proteínas dos organismos são profundamente influenciadas por pequenas alterações no pH sanguíneo (MACARI, 1994). O equilíbrio ácido-base está diretamente ligado aos eletrólitos ingeridos pelas aves. O balanço eletrolítico (BE) pode influenciar o crescimento, o apetite, o desenvolvimento ósseo, a resposta ao estresse térmico e o metabolismo de certos nutrientes, como aminoácidos, minerais e vitaminas (PATIENCE, 1990). Os principais elementos envolvidos nesse equilíbrio são os cátions sódio (Na+), potássio (K+) e magnésio (Mg++), os ânions cloro (Cl- ), bicarbonato (HCO3 - ) e fosfato dibásico (H2 PO4 - ), além de algumas proteínas. O Na+ e o Cl- contribuem, sobretudo para a pressão osmótica do plasma, enquanto o Mg++, os fosfatos e as proteínas, para pressão osmótica do fluido intracelular (GONZÁLEZ & SILVA, 1999). Mongin (1981) estudou os fundamentos do balanço eletrolítico (BE) para aves e suínos e descreveu o equilíbrio entre os íons por um cálculo envolvendo os principais deles, cuja fórmula representativa é o resultado da soma dos íons Na+ e K+ subtraindo-se o íon negativo Cl-, denominado BE. Porém Araújo (2011) propôs um novo cálculo de balanço eletrolítico onde os eletrólitos que não são contabilizados pela equação de Mogin (1981) fossem então contabilizados para então saber influência de seus mEq sobre o metabolismo das aves.Materiais e Métodos
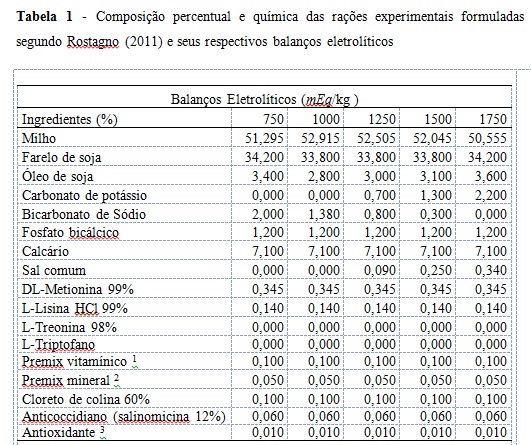
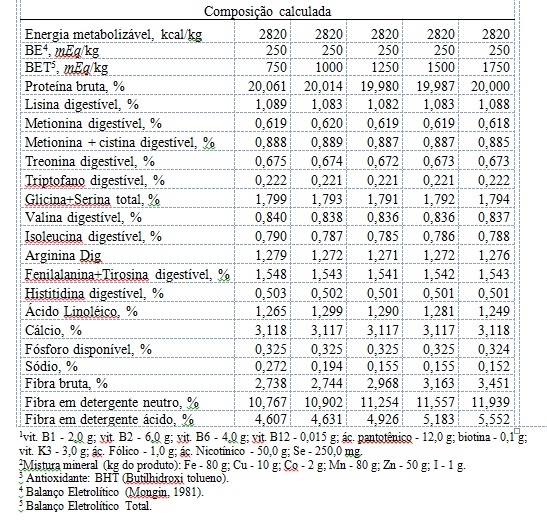
Os experimentos foram conduzidos no setor de avicultura do Instituto Federal Norte de Minas Gerais, em Januária, Minas Gerais, no período de agosto de 2016 a dezembro de 2016. A fim de atingir os objetivos propostos, foram confeccionadas cinco dietas, de mesmo valor de balanço eletrolítico (200 mEq/kg) segundo Mongin (1981), de acordo com a equação 1: Equação 1: BE, mEq/kg da ração = (mg/kg de Na+ da ração/22,990) + (mg/kg de K+ da ração /39,102 ) - (mg/kg de Cl- da ração/35,453) De posse da equação proposta por Araújo et al. (2011) as dietas possuiram 750, 1000, 1250, 1500, 1750 mEq/kg respectivamente (tabela 1). Calculo de balanço eletrolítico total (BET) segundo Araújo et al. (2011) de acordo com a equação 2: Equação 2: BET, mEq/kg da ração = [(mEq+/kg Ca2++mEq+/kg K+ + mEq+/kg Na++ mEq+/kg Mg2+)+ (mEq+/kg Lys + mEq+/kg Met + mEq+/kg Tre + mEq+/kg Trp + mEq+/kg Val + mEq+/kg Glu + mEq+/kg Chol)] – [(mEq-/kg PO43- + mEq-/kg SO4-2 + mEq-/kg Cl- + mEq-/kg HCO3-) + (mEq-/kg Lys + mEq-/kg Met + mEq-/kg Tre + mEq-/kg Trp + mEq-/kg Val + mEq-/kg Glu + mEq-/kg Chol)] Foram utilizadas 240 codornas japonesas, durante o período de 25 a 85 dias de vida, distribuído em um delineamento inteiramente casualizado, num total de cinco tratamentos (750, 1000, 1250, 1500, 1750 mEq/kg), oito repetições e seis aves por unidade experimental. As aves foram alojadas em gaiolas de arame galvanizado, com dimensões de 1,0m de comprimento x 0,25 m de largura x 0,20 m de altura, dispostas em um (1) andar, emparelhadas de duas a duas. Cada gaiola foi subdividida em quatro repartições iguais de 0,25 m. As laterais e o piso das gaiolas foram confeccionados com arame galvanizado. O comedouro e o bebedouro utilizados foram do tipo calha, em chapa metálica galvanizada, e ambos colocados percorrendo a extensão das gaiolas, sendo o comedouro na parte frontal e o bebedouro na parte posterior da gaiola. As rações experimentais foram formuladas para atender ou exceder as exigências nutricionais das codornas e valendo-se de informações sobre as composições dos ingredientes apresentadas em (ROSTAGNO et al. 2011). As rações foram fornecidas à vontade, duas vezes ao dia, às 8:00 e às 16:00 h. As rações experimentais foram isocalóricas, isoaminoacídicas para metionina + cistina, lisina e treonina digestíveis, e isofosfóricas. Os tratamentos consistiram de rações com os seguintes balanços eletrolíticos: 750, 1000, 1250, 1500 e 1750 (mEq/kg) (Tabela 1). Todas as dietas possuirão o mesmo balanço eletrolítico (250 mEq/kg) reportado por (MONGIN 1981). Os bebedouros foram limpos diariamente pela manhã. O programa de luz utilizado foi o de 17 (dezessete) horas de luz, por meio de um controlador de luz do tipo “timer”. As mensurações de temperatura e umidade dentro do galpão foram registradas por meio de termômetros de máxima e mínima e de bulbo seco e bulbo úmido. Foram analisadas as seguintes variáveis de produção: postura (%), peso médio dos ovos (g), consumo de ração (g/ave/dia), conversão alimentar por massa de ovos (g de ração/g de ovo), conversão alimentar por dúzia de ovos (g de ração/dz de ovo) e uniformidade (%). Para o controle do consumo as rações de cada repetição foram acondicionadas em baldes plásticos, devidamente identificados. O consumo de ração foi medido ao término de cada período de 28 dias, por meio da diferença entre a ração fornecida e a sobra. Também, tomou-se o cuidado de pesar as aves mortas e as sobras das rações, para realizar o controle do consumo, do ganho de peso dos animais, da postura e da conversão alimentar ao término de cada período. A coleta dos ovos foi feita diariamente às 16:00 h e a produção de ovos foi obtida em porcentagem ave/dia. Para obtenção dos dados de peso médio dos ovos, foi coletada, ao acaso, nos quatro últimos dias de cada período, uma amostra de seis ovos/parcela/dia, sendo estes pesados em balança com precisão de 0,01g. Após a pesagem dos ovos, estes foram quebrados e suas cascas mantidas identificadas para serem secas e pesadas. Para obtenção da massa de ovos, tomou-se a produção de ovos postos/ ave/ dia, multiplicando-se pelo peso médio dos ovos. A conversão alimentar foi obtida de duas maneiras diferentes: dividindo-se o consumo médio diário de ração pela produção média diária em dúzias de ovos (conversão g/dz), e dividindo-se o consumo médio de ração pela produção média de ovos em g (conversão kg/kg). Para a variáveis qualitativas foram avaliadas: densidade, coloração da gema, unidade Haugh, peso da gema, peso do albúmen, peso da casca e peso do ovo utilizando-se seis ovos de cada repetição, coletados aleatória e diariamente do total de ovos coletados nos três últimos dias de cada período. Foram feitas imersões dos ovos em diversas soluções salinas com densidades conhecidas com os devidos ajustes. Os ovos foram colocados nos recipientes com as soluções, da menor para a maior densidade, e retirados ao flutuarem registrando-se os valores das densidades correspondentes às soluções dos recipientes. A pigmentação foi obtida através do leque calorimétrico ROCHE, numa escala de valores de 1 a 15 (do amarelo claro à cor de abóbora respectivamente). Os ovos de cada repetição e de cada dia foram pesados individualmente em balança com precisão de 0,001 g e, depois de pesados, foram identificados e quebrados. A gema de cada ovo foi pesada e a respectiva casca lavada e seca ao ar para posterior obtenção do peso da casca sem a membrana interna. O peso do albúmen foi calculado como a diferença entre o peso do ovo e os pesos da gema e da casca. A avaliação estatística foi feita utilizando-se análise de covariância pelo procedimento PROCGLM (SAS, 2002) onde o peso inicial das aves foi considerado como covariável. Após a comprovação de diferenças estatísticas equações de regressão foram delineadas utilizando o procedimento PROCREG (SAS, 2002). A uniformidade foi avaliada através do método da análise percentual.Resultados e Discussão
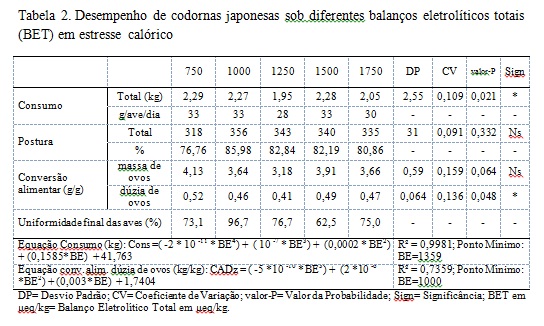
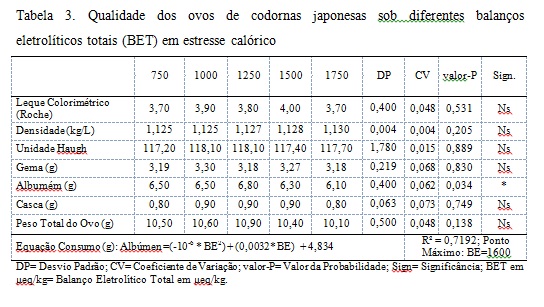
Durante o experimento as médias de temperatura máxima e mínima foram respectivamente 36,1ºC e 23,8 ºC e umidade relativa do ar de 48,96%, o suficiente para induzir o estresse calórico nas aves. Os parâmetros de postura, conversão alimentar por massa de ovos não foram afetados pelos balanços eletrolíticos totais (BET) experimentais (Tab. 2, P>0,05). Com base nos parâmetros de postura, estes estão coerentes com os citados por Moraes (2010), onde trabalhando com balanço eletrolítico para codornas japonesas na fase de produção, também não encontrou efeito significativo com relação aos balanços eletrolíticos (BE) estudados. Para conversão alimentar por massa de ovos, esses resultados se assemelham aos encontrados por e Bezerra et al. (2011) que não verificaram efeito de significância para esse parâmetro em seus trabalhos. Em contrapartida, Moraes (2010) relatou efeito significativo (P<0,05) com relação ao balanço eletrolítico, mostrando que quanto maior a inclusão de NaHCO3 na dieta, melhor é a conversão alimentar. Estes estudos apontados consideraram apenas uma pequena parte dos eletrólitos da dieta utilizando o cálculo clássico de BE (MONGIN, 1981). Talvez por considerarem uma pequena parte dos eletrólitos da dieta, cargas não consideradas no cálculo clássico de BE possam interferir no resultado de desempenho das aves, gerando uma grande variabilidade de resultados contrastantes entre si. O cálculo do BET é novo e possui poucos trabalhos relatados ainda. À medida que mais estudos forem gerados maior será o conhecimento a respeito do equilíbrio ácido-base e suas respostas sobre a performance dos animais. Os valores de consumo e conversão alimentar por dúzia de ovos foram afetados pelos diferentes BET (P<0,05) e também os valores de uniformidade. A conversão alimentar por dúzia de ovos e o consumo podem ser representados pelas equações presentes na tabela 2. Os valores de BE que propiciam a conversão alimentar por dúzia de ovos mínima e melhor uniformidade é BET = 1000 em μeq/kg de cargas nas rações. O valor de BE que propicia o menor consumo sob mesmo desempenho, descrita segundo a equação, é o BET = 1359 μeq/kg. Resultados semelhantes aos encontrados neste trabalho foram observados por Moraes (2010) onde foi encontrado um efeito significativo para o consumo de ração, com uma dieta de 320 mEq/kg contendo maiores níveis de carbonato de potássio e bicarbonato de sódio e menores níveis de sal comum, resultando em menor consumo de ração. Este autor inferiu que essa redução de consumo pode estar relacionada ao excesso de eletrólitos e/ou as propriedades químicas do carbonato de potássio. Raquel et al. (2010) não observaram efeito significativo trabalhando com níveis de cloro para codornas italianas destinadas à produção de carne, sendo os níveis de cloro com 0,07 a 0,32% de cloro na ração a base de milho e farelo de soja contendo 0,15 de sódio e 0,94% de potássio. Uma das estratégias a fim de se diminuir a carga de eletrólitos na dieta seria a substituição do NaCl pelo NaHCO3 como fonte de sódio. Através desta manipulação consegue-se manipular erroneamente a quantidade de íons negativos da dieta através da retirada de Cl- utilizando a equação de BE proposta por Mongin (1981). Entretanto sabe-se que os íons bicarbonato também possuem a capacidade de interferir no equilíbrio ácido-base fisiológico, explicando a falta de resultados reportados por Raquel et al. (2010). Vieira et al. (2015) onde também não observaram efeito significativo em sua pesquisa sobre os níveis do balanço eletrolítico e proteína bruta sob as variáveis de desempenho para codornas japonesas utilizando o cálculo proposto por Mongin (1981). Com relação à conversão alimentar por dúzia de ovos, no qual houve efeito significativo (P<0,05) difere dos resultados obtidos por Vieira et al. (2015) estudando os níveis de balanço eletrolítico e proteína bruta para codornas japonesas na fase de postura. Os resultados reportados neste trabalho demonstram que outros íons presentes nas dietas (que não apenas Na, Cl e K) são capazes de influenciar a performance das aves, e a equação de Mogin (1981) subestima a influência dos eletrólitos sob a fisiologia animal. Em situações de estresse calórico a perda de íons bicabornato pelos rins das aves é potencializada pela alcalose respiratória, sendo, portanto a manipulação do BET da dieta uma estratégia para minimizar esses efeitos deletérios ao metabolismo das codornas poedeiras. Sob o aspecto da zootecnia de precisão atributos como bem estar animal, comportamento de estresse, e nutrição de precisão são considerados dentro da matriz de parâmetros produtivos a serem avaliados (SEVEGNANI et al., 2005). Dentro de um conceito de nutrição de precisão a equação de BET demonstra maior capacidade de quantificar os eletrólitos responsáveis por alterações metabólicas nos animais que o método clássico de cálculo de BE proposto por Mongin (1981). Também foram avaliados parâmetros relacionados a qualidade do ovo. Os valores de densidade, coloração da gema, Unidade Haugh, peso da gema, peso da casca e peso do ovo não foram afetados pelos balanços eletrolíticos totais (BET) experimentais (Tab. 3, P>0,05). Para a unidade Haugh, não foram encontrados efeito significativo (P>0,05). Bezerra et al. (2011), encontraram valores entre 88 e 90, bem abaixo dos obtidos nesse trabalho. Para os parâmetros peso de gema e densidade do ovo Neves (2015) estudando a manipulação de eletrólitos na dieta de poedeiras em fase de declínio de postura em condições de altas temperaturas, não constatou efeito significativo (P>0,05) sobre os BE estudados, porém, este observou efeito significativo (P<0,05) do ciclo sobre todos os parâmetros de qualidade do ovo, pois a temperatura durante cada ciclo avaliado sofreu uma amplitude em média de 18,6ºC, assim este ressalta ainda que quanto maior a porcentagem de casca, nesse caso para o primeiro ciclo, melhor os índices de espessura de casca e densidade. Em relação ao peso dos ovos, Bezerra et al. (2011) encontraram valores entre 10,0 e 10,3 g com o balanço eletrolítico da ração variando de 174,18 a 245,44 mEq/kg. O tamanho da gema é afetado, nutricionalmente, em função dos níveis de energia e metionina da dieta (BRUMANO et al., 2010). Dificilmente se evidencia a variação deste parâmetro em função da manipulação do BE. Apesar de se esperar melhorias na qualidade da casca dos ovos com a manipulação do BET este fato não foi verificado. Entretanto futuros estudos deverão ser conduzidos com uma variação de maior amplitude do BET a fim de se evidenciar a influência sobre esse parâmetro em específico. Os valores de peso do albúmen foram afetados pelos diferentes BE (P<0,05). O peso do albúmen pode ser representado pela equação presente na tabela 3. O valor de peso do albúmen máximo descrito segundo a equação é o valor de BE= 1600 em μeq/kg de cargas nas rações. Esses resultados são coerentes aos encontrados por Moraes et al. (2010) onde os maiores pesos foram encontrados nos tratamentos que continham 196 e 241 mEq/kg de BE. Bezerra et al. (2011) trabalhando com níveis de BE de 174,18 até 245,44 mEq/kg, relatam que entre esses valores não houve influência no peso do albúmen.. No caso de frangos de corte as situações adversas, envolvendo altas temperaturas, acabam por desencadear alcalose respiratória resultando assim queda no desempenho (ALDRIGUI, 2014). Os resultados encontrados nesse trabalho podem ter tido influência do estresse calórico sofrido pelas aves devido à alta temperatura e baixa umidade relativa do ar obtendo então efeito significativo (P<0,05). A manipulação do BE melhora a retenção hídrica dos animais, principalmente em situações de estresse calórico (BEKER e TEETER, 1994). A albumina (presente no albúmen) apresenta grande retenção de moléculas de água, portanto a retenção de água no organismo da ave e consequente disponibilidade para a irrigação sanguínea do trato reprodutivo (oviduto) tem influência direta sobre o peso do albúmen (MACARI e FURLAN, 2002). Com base nestes fatos a manipulação do BET da dieta pode ter propiciado maior retenção hídrica no organismo das aves resultando em maior peso do albúmen dos ovos.Conclusões
Os diferentes balanços eletrolíticos totais (BET) são capazes de influenciar os parâmetros de desempenho e qualidade dos ovos. O valor de BET que propiciou menor consumo sob mesmo desempenho foi de BET1=1359 μeq/kg, e o que resultou em melhor conversão alimentar por dúzia de ovos e uniformidade é de BET = 1000 μeq/kg. Para a qualidade do ovo o BET que propicia peso máximo do albúmen é de BET= 1600 μeq/kg.Gráficos e Tabelas