![]()
COMPONENTES BIOQUIMICOS EM SORGO COM DOSE DE SILICIO E ESTRESSE HÍDRICO AVALIADOS ATRAVÉS DA MULTIVARIADA
2 - Graduanda do curso de Zootecnia da Universidade Federal Rural da Amazônia - UFRA
3 - Graduanda do curso de Agronomia da Universidade Federal Rural da Amazônia – UFRA
4 - Graduanda do curso de Agronomia da Universidade Federal Rural da Amazônia – UFRA
5 - Graduanda do curso de Agronomia da Universidade Federal Rural da Amazônia – UFRA
6 - Graduanda do curso de Agronomia da Universidade Federal Rural da Amazônia – UFRAGraduanda do curso de Agronomia da Universidade Federal Rural da Amazônia – UFRA
7 - Doutora em Solo da Universidade Estadual Paulista – UNESP.
8 - Professor Adjunto da Universidade Federal Rural da Amazônia – UFRA
RESUMO -
O sorgo é uma cultura de importância no Brasil. O estresse hídrico pode induzir variadas respostas bioquímicas e fisiológicas em plantas e o silício é um micromineral que pode agir como atenuador desse estresse as plantas. Portanto, o objetivo do trabalho foi avaliar os componentes bioquímicos na raiz e na folha em plantas de sorgo forrageiro em condições de estresse hídrico e em diferentes concentrações de silício, através da análise multivariada. O experimento foi em casa de vegetação da Universidade Federal Rural da Amazônia (UFRA), utilizando plantas de sorgo (Sorghum bicolor [Moench.]). O delineamento experimental foi inteiramente casualizado (DIC) com o fatorial 2×4 com 7 repetições, referente a duas condições hídricas (irrigado e déficit hídrico) e quatro doses de silício (Si) (0.5, 1.0, 1.5 e 2.0 µM). Há distinção na quantidade de componentes bioquímicos nas raízes e folhas de plantas de sorgo. E esta quantidade também varia de acordo as condições de estresse hídrico no solo.
BIOCHEMICAL COMPONENTS IN SORGHUM WITH DOSES OF SILICON AND WATER STRESS ASSESSED THROUGH THE MULTIVARIATE
ABSTRACT - Sorghum is a culture of importance in Brazil. Water stress can induce varied biochemical and physiological responses in plants and silicon is a micromineral that can act as attenuator of this stress to plants. The objective of this work was to evaluate the biochemical components in root and leaf in forage sorghum plants under conditions of water stress and in different concentrations of silicon, through the multivariate analysis. The experiment was carried out in a greenhouse at the Federal Rural University of Amazonia (UFRA), using sorghum plants (Sorghum bicolor [Moench.]). The experimental design was completely randomized (DIC) with factorial 2x4 with 7 replicates, referring to two water conditions (irrigation and water deficit) and four silicon concentrations (Si) (0.5, 1.0, 1.5 and 2.0 μM). There is a distinction in the amount of biochemical components in the roots and leaves of sorghum plants. And this amount also varies according to the conditions of soil water stress.Introdução
Aproximadamente entre 10-15% da área cultivada com sorgo no Brasil é utilizado como ensilagem e tem-se destacado devido à alta produção (AVELINO et al., 2011). O estresse hídrico pode induzir variadas respostas bioquímicas e fisiológicas em plantas, sendo um fator de grande influência para o crescimento e produção vegetal (NEMANI et al., 2003). O silício é um micromineral de utilização recente como atenuador de estresses abióticos, dentre eles o estresse hídrico (SOUZA et al., 2014). Portanto, o objetivo do trabalho foi avaliar os componentes bioquímicos na raiz e na folha em plantas de sorgo forrageiro em condições de estresse hídrico e em diferentes concentrações de silício, através da análise multivariada.Revisão Bibliográfica
A condição de insuficiência de água é considerada uma das maiores limitações agrícola no mundo, ocasionando redução do conteúdo relativo de água, atividade da redutase do nitrato, proteínas, amônio e nitrato nos tecidos, e pela elevação das concentrações de aminoácidos (SOUZA et al., 2014). O silício é um micromineral de utilização recente como atenuador de estresses abióticos, dentre eles o estresse hídrico. Porém, há escassez de trabalhos científicos relacionados ao comportamento do sorgo sobre deficiência hídrica e nas diferentes concentrações de silício para produção de forragem, objetivando à determinação de parâmetros de resposta baseados nas modificações bioquímicas dessa cultura e nas relações hídricas do sistema solo-planta (SOUZA et al., 2014).Materiais e Métodos
O experimento foi em casa de vegetação da Universidade Federal Rural da Amazônia (UFRA), campus Capitão Poço, localizado nas Coordenadas: Latitude 01º44'47" Sul; Longitude 47º03'34" Oeste. Utilizou-se plantas de sorgo (Sorghum bicolor [Moench.]) forrageiro da variedade BR 700. O delineamento experimental foi inteiramente casualizado (DIC) com o fatorial 2x4 com 7 repetições, referente a duas condições hídricas (irrigado e déficit hídrico) e quatro doses de silício (Si) (0.5, 1.0, 1.5 e 2.0 µM) que foram avaliados na posição das raízes (R) e folhas das plantas de sorgo (ER-Si e EF-Si). As plantas foram irrigadas duas vezes ao dia (manhã e tarde) com solução nutritiva de Hoagland & Arnon (1950). Analisou-se nas folhas e raízes o conteúdo relativo de água pelo método descrito por Slavick (1979). A quantidade de amônio livre pelo método de Weatherburn (1967). A concentração de nitrato pelo método de Cataldo et al. (1975). A atividade da redutase do nitrato pelo método Hageman e Hucklesb (1971), para obtenção das proteínas solúveis totais foi pelo método de Bradford (1976), e para mensurar os aminoácidos solúveis totais a metodologia preconizada por Peoples et al. (1989). Para a análise de agrupamento, uma matriz de semelhança foi construída com a distância euclidiana e a ligação dos grupos foi realizada com o método de Ward (SNEATH; SOKAL, 1973). As análises foram conduzidas no programa STATISTICA 7.0 (StatSoft. Inc., Tulsa, OK, USA). Simultaneamente às análises estatísticas, as pressuposições básicas da análise de variância, normalidade dos erros e homogeneidade das variâncias foram testadas para todas as variáveis avaliadas (dados não apresentados).Resultados e Discussão
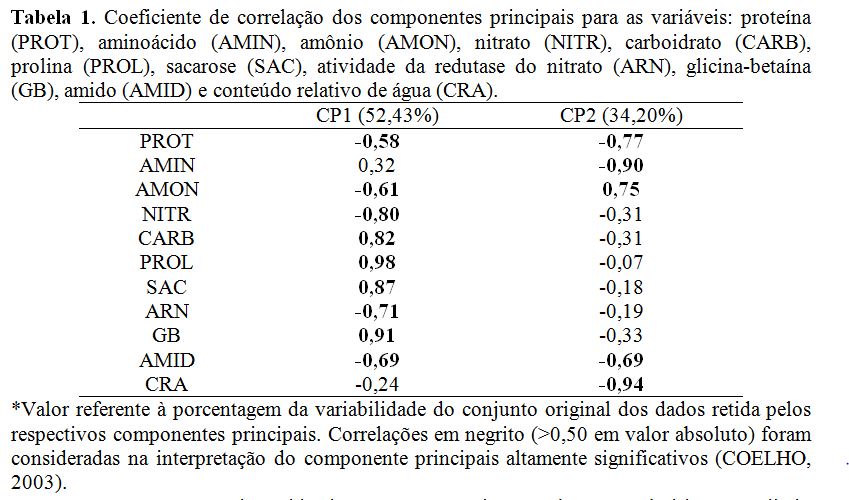
Na análise de componentes principais (ACP) ocorreu a formação de um plano bidimensional (Figura 1) com dois componentes principais CP1 e CP2, correspondendo a 86,63 % da informação original: 52,43 e 34,20 %; respectivamente no CP1 e CP2. Este resultado condiz com o critério estabelecido por Sneath e Sokal (1973). Na CP1 por ordem de importância, os atributos que apresentaram coeficientes de correlação positivos foram a PROL (0,98), GB (0,91), SAC (0,87) e CARB (0,82). Enquanto, os negativos ARN (-0,71), AMON (-0,61), NITR (-0,80), AMID (-0,69) e PROT (-0,58). As correlações são apresentadas na Tabela 1 e representadas pelas setas de cada atributo e sua projeção na Figura 1. Na CP2 ficou retida menor quantidade de variáveis. A PROT e AMID também apresentaram coeficientes significativos, assim como na CP1. As variáveis que apresentaram por ordem de importância maiores coeficientes de correlação positivos AMON (0,75) e negativos CRA (-0,94) AMIN (-0,90), PROT (-0,77), AMID (0,69). Na planta ocorre um aumento da quantidade de PROL, GB, SAC e CARB, enquanto os teores de ARN, AMON, NITR, AMID e PROT decrescem. Em plantas de milho em deficiência hídrica ocorreu a redução de ARN, CRA, PROT, NIT, AMON e aumento de AMIN devido a deficiência hídrica porque favorece a degradação das proteínas pelas enzimas proteolíticas e aumenta os teores de AMIN na planta (SOUZA et al., 2014). As correlações positivas nos parâmetros carboidratos e sacarose na CP1 (Tabela 1 e Figura 1) no sistema radicular é uma resposta de tolerância ao estresse hídrico, ocasionando ajustamento osmótico no metabolismo da planta e reduzindo seus potenciais osmóticos pelas diferenças de gradientes de concentrações mantendo a turgescência e consequentemente retardando a desidratação nos tecidos das plantas (SALISBURY; ROSS,2012). As concentrações de amido decresceram nas plantas sob estresse hídrico sem silício possivelmente por causa da ação principalmente pelas enzimas α e β amilase convertendo amido em carboidratos e sacarose. A prolina e glicina betaína também apresentaram correlações positivas (Figura 1) devido à quebra das proteínas por enzimas específicas convertendo em aminoácidos. Pois, condição de estresse hídrico aumenta a atividade de enzimas proteolíticas, que quebram as proteínas de reserva e também da diminuição da síntese “de novo” das mesmas (OLIVEIRA NETO, 2010). Os coeficientes de correlação negativo no parâmetro nitrato na CP1 (Tabela 1 e Figura 1) devem-se ao baixo conteúdo de água no solo, pois o nitrato (NO3-) é a principal forma de nitrogênio absorvido do solo pelas plantas, seguido do amônio (NH4+) (MALAVOLTA, 2006). No entanto, quando o NO3- encontra-se em quantidades baixas dificulta o processo de absorção pelas raízes e consequentemente diminui o transporte deste para a parte aérea via xilema (SHANER; BOYER, 1976), diminuindo a atividade da redutase do nitrato, essa redução também foi nesse experimento com correlação negativa (Tabela 1 e Figura 1).Conclusões
Há distinção na quantidade de componentes bioquímicos nas raízes e folhas de plantas de sorgo. E esta quantidade também varia de acordo as condições de estresse hídrico no solo.Gráficos e Tabelas