![]()
Teores de Prolina, Clorofila e Carotenoides Em Pueraria phaseoloide Sob Estresse Hídrica
2 - Mestre em Zootecnia, Universidade Estadual do Sudoeste da Bahia – UESB
3 - Professora da Universidade Estadual do Sudoeste da Bahia – UESB, Campus de Itapetinga, BA
4 - Graduando (a) em Ciências Biológicas, Universidade Estadual do Sudoeste da Bahia – UESB, Campus de Itapetinga, BA
5 - Graduando (a) em Ciências Biológicas, Universidade Estadual do Sudoeste da Bahia – UESB, Campus de Itapetinga, BA
6 - Graduando (a) em Zootecnia, Universidade Estadual do Sudoeste da Bahia – UESB, Campus de Itapetinga, BA
7 - Graduando (a) em Zootecnia, Universidade Estadual do Sudoeste da Bahia – UESB, Campus de Itapetinga, BA
8 - Pós-doutoranda em Zootecnia, Universidade Estadual do Sudoeste da Bahia – UESB, Campus de Itapetinga, BA
RESUMO -
Objetivou-se avaliar os teores de prolina, clorofila e carotenoides em Pueraria phaseoloide sob condições de deficiência hídrica e adubação nitrogenada. O experimento foi realizado em casa de vegetação, UESB, Itapetinga, BA. O ensaio foi conduzido em esquema fatorial 4 x 2, sendo quatro regimes hídricos (25, 50 75 e 100% da capacidade de campo) e duas doses de nitrogênio (0 e 75 kg de N/ha), disposto em DIC, com quatro repetições. Ao final do período de estresse foram realizadas as análises, para as quais foram coletadas folhas completamente expandidas e utilizadas frescas para clorofilas e carotenoides e secas para prolina. Para a extração da prolina utilizou-se ácido sulfosalicílico 3% e para as clorofilas e carotenóides, o Dimetilsulfóxido. A interação entre doses de nitrogênio e regime hídrico foi verificada a 5% de probabilidade e o regime hídrico foi avaliado por análise de regressão e as doses de nitrogênio pelo teste F. Houve efeito quadrático (P<0,05) dos regimes hídricos sobre os teores de prolina, chegando a aumentar cerca de 5 vezes mais no regime hídrico de 25% CC quando comparado com os demais. Os teores de clorofilas e carotenoides não apresentaram alterações em função dos regimes hídricos e somente a razão clorofila a/b aumentou em presença da adubação nitrogenada. O acúmulo de prolina ocorre em P. phaseoloide em regimes hídricos mais baixos, o que indica uma boa tolerância dessas plantas ao estresse hídrico. Isso é reforçado, uma vez que, as clorofilas e carotenoides não são afetados pela deficiência hídrica.
Prolina, Chlorophila and Carotenoids Contents In Pueraria Phaseoloide Under Water Stress
ABSTRACT - The objective of this study was to evaluate the proline, chlorophyll and carotenoids contents in Pueraria phaseoloide under conditions of water deficit and nitrogen fertilization. The experiment was carried out in a greenhouse, UESB, Itapetinga, BA. The experiment was conducted in a 4 x 2 factorial scheme, with four water conditions (25, 50, 75 and 100% of the field capacity) and two doses of nitrogen (0 and 75 kg of N/ha) disposed in DIC, with four replicates. At the end of the stress period, the analyzes were performed, for which completely expanded leaves were collected and used fresh for chlorophyll and carotenoids and dried for proline. For proline extraction was used 3% sulfosalicylic acid and for chlorophylls and carotenoids, Dimethyl sulfoxide. The interaction between nitrogen doses and water conditions was verified at 5% probability and the water regime was evaluated by regression analysis and nitrogen doses by the F test. There was a quadratic effect (P<0.05) of the water conditions on the proline content, reaching about 5 times higher in the water regime of 25% CC when compared to the others. The levels of chlorophyll and carotenoids did not change as a function of water conditions and only the chlorophyll a/b ratio increased in the presence of nitrogen fertilization. The accumulation of proline occurs in P. phaseoloide at lower water regimes, which indicates a good tolerance of these plants to water stress. This is reinforced, since, chlorophylls and carotenoids are not affected by water deficiency.Introdução
No mundo globalizado, diante de uma pecuária competitiva, faz-se necessária a intensificação dos sistemas de produção animal no Brasil. Além de produtivos, os sistemas devem ser sustentáveis e que sejam minimizados os prejuízos ao meio ambiente. Uma alternativa bastante promissora é o uso de leguminosas em sistemas consorciados com gramíneas, que pode contribuir para reduzir os problemas decorrentes da degradação dos ecossistemas.
O conhecimento sobre as leguminosas se torna importante devido aos problemas encontrados na sua utilização em sistemas consorciados, como a baixa persistência. Dessa forma, o estudo do crescimento e desenvolvimento para sua implantação em pastagens se torna essencial.
Além disso, sendo o nitrogênio o nutriente de maior importância para o crescimento e a manutenção das plantas, a utilização no estabelecimento de leguminosas pode favorecer a sua persistência, sem interferir na fixação biológica.
A verificação das características de tolerância à deficiência hídrica de leguminosas tropicais aliada à adubação nitrogenada poderá contribuir para o conhecimento mais expressivo de adaptação de plantas sob condições de estresse. Nesse sentido, as leguminosas, além de serem uma opção como alimentação animal, poderão contribuir favoravelmente minimizando danos ao solo decorrente da atividade pecuária.
Objetivou-se avaliar os teores de prolina, clorofila e carotenoides em Pueraria phaseoloides sob condições de deficiência hídrica e adubação nitrogenada.
Revisão Bibliográfica
A Pueraria phaseoloides é uma leguminosa herbácea perene e tem sua origem no sudoeste da Ásia e foi introduzida no Brasil a partir da década de 1940, com o objetivo de ser utilizada como cobertura de solo nos plantios de seringueira na Amazônia (Valentim, 2010). Desde então sua utilização é, principalmente, na cobertura do solo em plantações de espécies perenes, como adubo verde, forrageira em pastos exclusivos ou consorciadas com gramíneas (Rasmo et al., 2010).
Conforme Valentim et al. (1984), esta leguminosa constitui-se em excelente fonte de proteína para o gado, além da sua relativa eficiência na fixação do nitrogênio atmosférico, melhorando consequentemente a fertilidade do solo.
O nitrogênio é o nutriente requerido em maior quantidade pelas plantas, por ser um constituinte das clorofilas, ácidos nucléicos e proteínas, moléculas fundamentais para todos os processos biológicos.
Conforme Costa et al. (2006), a deficiência de nitrogênio tem sido apontada como a principal causa para a redução da produtividade e degradação das pastagens, que ocorre em áreas que não receberam adubação nitrogenada ou que receberam em baixos níveis. Sua deficiência reduz a taxa fotossintética, proporcionando um crescimento reduzido nas plantas e se fornecido adequadamente, em condições favoráveis, proporciona aumento na produção de matéria seca.
A deficiência hídrica é outro fator que interfere na produção das plantas e essa condição ambiental é comum em regiões do nordeste brasileiro. O conteúdo total de clorofilas nas folhas pode ser um indicativo de estresses hídrico e salino, uma vez que já foram detectadas reduções significativas desses pigmentos em várias espécies de plantas (Gratani & Varone, 2004; Chylinski et al., 2007).
A prolina, ativamente acumulados em resposta a estresses abióticos, sobretudo, hídrico e salino, resulta em aumento da osmolaridade celular, promovendo uma manutenção no turgor, que é imprescindível à expansão celular. Assim, é conhecida como agente osmorregulador em muitas espécies de plantas, aumentando várias vezes com a redução do potencial hídrico da folha (Reddy et al., 2004; Ashraf & Harris, 2004).
O conhecimento das respostas da planta forrageira em condições de estresse hídrico é de grande importância para auxiliar no entendimento dos efeitos do período seco na produção de forragem, possibilitando o uso de práticas de manejo para melhor utilização do pasto durante esses períodos (Araújo et al., 2010).
Materiais e Métodos
O experimento foi realizado em casa de vegetação, UESB, Itapetinga-BA, em um esquema fatorial 4 x 2, sendo quatro regimes hídricos (25, 50 75 e 100% da capacidade de campo (CC)) e duas doses de nitrogênio (0 e 75 kg de N/ha), disposto em DIC, com quatro repetições.
As mudas de Pueraria phaseoloides foram produzidas a partir de sementes, mantendo-se 4 plantas por vaso. A adubação nitrogenada (75kg N/ha) foi realizada em dose única, após 30 dias da semeadura.
Para calcular a reposição de água para cada regime hídrico foi determinada a máxima capacidade de retenção de água do solo por meio da diferença de peso dos vasos com solo seco e molhado (após encharcar e escoar a água), a qual foi em torno de 18%. Assim, diariamente, por meio de pesagem, a água foi completamente reposta nos vasos com solo próximo à capacidade de campo e proporcionalmente a 75, 50 e 25% do peso nos outros regimes hídricos.
Após 28 dias de estresse, foram coletadas folhas completamente expandidas, as quais foram utilizadas frescas para as análises de clorofilas e carotenoides e secas para a prolina.
Para a extração da prolina, a massa seca de folhas foi homogeneizada em ácido sulfosalicílico 3%, sendo mantido sob agitação constante à temperatura ambiente, por 60 minutos. Em seguida, o material foi filtrado e a quantificação realizada segundo Bates (1973).
Para a extração das clorofilas e carotenóides, fragmentos de folhas coletadas (10:00 horas) foram, imediatamente, pesados e submetidos à extração com Dimetilsulfóxido por 72 horas. A quantificação foi feita de acordo com Wellburn (1994). Os resultados foram submetidos à análise de variância, considerando como fontes de variação a dose de nitrogênio, o regime hídrico e a interação dose de nitrogênio × regime hídrico, testados a 5% de probabilidade. A interação foi desdobrada, ou não, de acordo com a significância, sendo o efeito do regime hídrico avaliado por análise de regressão e das doses de nitrogênio pelo teste F.
Resultados e Discussão
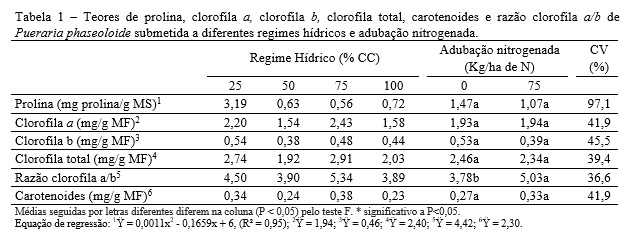
Não houve interação significativa para os teores de prolina, clorofila a, clorofila b, clorofila total, carotenoides e razão clorofila a/b em Pueraria phaseoloide (Tabela 1).
O teor de prolina apresentou efeito quadrático (P<0,05) do regime hídrico. No regime hídrico de 25% CC, o teor de prolina foi cerca de 5 vezes maior quando comparado com os demais. Segundo Maia et al. (2007), o acúmulo de prolina em tecidos foliares de diversas espécies quando submetidos às condições de estresse hídrico tem variações genotípicas. O aumento nos teores de prolina em P. phaseoloide demonstrara sua potencialidade genética em aumentar a concentração desse aminoácido em função do estresse hídrico. Ao mesmo tempo, nos regimes hídricos de 50 e 75% CC não houve aumento desse aminoácido, indicando que, nesses casos, as plantas não estavam sob estresse.
O efeito quadrático da produção de prolina, com altas concentrações em 25% CC, demonstrou a capacidade de ajuste osmótico e uma possível adaptação favorável dessa planta em condições de baixa umidade no solo. A biossíntese e o acúmulo de solutos osmoticamente ativos, como o aminoácido prolina, no vacúolo ou no citosol, tem função osmoprotetora, o que mantém o equilíbrio hídrico e preserva a integridade celular de proteínas, enzimas e membranas (ABDUL JALEEL et al., 2007).
Os regimes hídricos não influenciaram (P>0,05) os teores de clorofila a, clorofila b, clorofila total e carotenoides, nem a razão clorofila a/b (Tabela 1). O nitrogênio apresentou efeito significativo (P<0,05) apenas para razão clorofila a/b, indicando menor quantidade de clorofila b quando se adubou com nitrogênio.
A clorofila não foi afetada pela deficiência hídrica, o que demonstra boa adaptação na manutenção desse pigmento em folhas intactas, porém, a redução no número de folhas em função do estresse reduz a área fotossintética mesmo sendo uma característica adaptativa na redução da perda de água. Segundo CARVALHO et al. (2003), decréscimos nos teores de clorofila podem ser sintoma característico de estresse oxidativo, e tem sido verificado em plantas sob estresse hídrico.
Conclusões
A Pueraria phaseoloide acumula prolina em estresse hídrico, porém os teores desse aminoácido aumentaram somente em regimes hídricos mais baixos, o que indica uma boa tolerância dessas plantas a esse estresse. Além disso, as clorofilas e carotenoides não são afetados pela deficiência hídrica, reforçando essas evidências.
Gráficos e Tabelas
Referências
ABDUL JALEEL, C.; MANIVANNAN, P.; KISHOREKUMAR, A.; SANKAR, B.; GOPI, R.; SOMASUNDARAM, R.; PANEERSELVAM, R. Alterations in osmoregulation, antioxidante enzymes and indole alkaloid levels in Catharanthus roseus exposed to water defi cit. Colloids and Surfaces B: Biointerfaces, v.59,v p.150-157, 2007.
ARAÚJO, S. A. C.; VASQUEZ, H. M.; CAMPOSTRINI, E.; NETTO, A. T.; DEMINICIS, B. B.; LIMA, E. S. Características fotossintéticas de genótipos de capim-elefante anão (Pennisetum purpureum Schum.), em estresse hídrico Acta Scientiarum. Animal Sciences, Maringá, v. 32, n. 1, p. 1-7, 2010.
ASHRAF, M.; HARRIS, P. J. C. Potential biochemical indicators of salinity tolerance in plants. Plant Science, Limerick, v.166, p.3-16, 2004.
BATES, L. S. Rapid determination of free proline for water-stress studies. Short Communication. Plant and Soil, v.39, n.1, p.205-207, 1973.
CARVALHO, L. M.; CASALI, V. W. D.; SOUZA, M. A.; CECON, P. R. Disponibilidade de água no solo e crescimento de Artemísia. Horticultura Brasileira, Brasília, v. 21, n. 4. p. 726-730, 2003.
CHYLINSKI, W. K.; LUKASZEWSKA, A. J.; KUTNIT, K. Drought response of two bedding plants. Acta Physiol Plant, v.29, p.399-406, 2007.
COSTA, K. A. P.; OLIVEIRA, I. P.; FAQUIN, V. Adubação Nitrogenada para Pastagens do Gênero Brachiaria em Solos do Cerrado. Santo Antônio de Goiás, GO. Embrapa (Documentos 192). 60 p, 2006.
GRATANI, L.; VARONE, L. Leaf key traits of Erica arborea L., Erica multiflora L. and Rosmarinus officinalis L. co-occuring in the Mediterranean maquis. Flora, v.199, p.58-69, 2004.
MAIA, P. S. P.; NETO, C. F. O.; CASTRO, D. S.; FREITAS, J. M. N.; LOBATO, A. K. S.; COSTA, R. C. L. C. Conteúdo relativo de água, teor de prolina e carboidratos solúveis totais em folhas de duas cultivares de milho submetidas a estresse hídrico. Revista Brasileira de Biociências, v.5, supl.2, p.918-920, 2007.
RASMO, A. K. B.; BARCELLOS, A. de O.; FENANDES, F.D. Plantas Forrageiras. Viçosa-MG: UFV, p.430-439, 2010.
REDDY, A. R.; CHAITANYA, K. V.; VIVEKANANDAN, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. Journal of Plant Physiology, v.161, n.11, p.1189-1202, 2004.
VALENTIM, J. F.; COSTA, A. L.; SILVA, C. S.; KOURE, J. Utilização de puerária na alimentação bovina. Comunicado técnico. Embrapa, Rio Branco - AC, 1984.
VALENTIM, J. F. Outras leguminosas forrageiras de importância econômica para a pecuária Brasileira. In: FONSECA, D.M.; MARTUSCELLO, J.A. (Eds.) Plantas forrageiras. Viçosa, MG: UFV, p.430-439, 2010.
WELLBURN, A. R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of diferente resolution. J. Plant Physiol., v. 144, n. 3, p. 307-313, 1994.
